In sum our work reveals that (m)RNA-clamped UAP56 not only promotes mRNA export by binding TREX-2 at the NPC. It can also be read out by the LENG8-PS module in PAXT to destine the RNA for decay. (9/x)
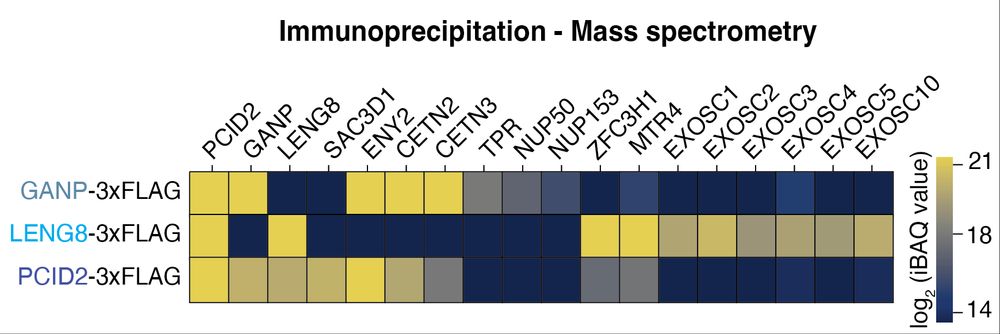
Perturbation of the LENG8–ZFC3H1 interface, or rapid depletion of either protein, leads to the upregulation of bona fide PAXT polyA-RNA targets. This demonstrates that the LENG8-PS module is functionally important for PAXT function. (8/x)
Alphafold reveals a conserved mode of interaction between LENG8 and the PAXT scaffolding subunit ZFC3H1, which we can validate in vivo. Thus, LENG8-PS constitutes a TREX-2-like module of PAXT. (7/x)
To our great surprise we found that LENG8-PS links to the ′Poly(A) tail exosome targeting (PAXT)’ connection, implicated in the degradation of polyadenylated nuclear RNA through the exosome. (6/x)
Turns out they do! Both additional complexes bind UAP56 in vitro, and, like TREX-2, stimulate the release of RNA from UAP56. Seems counter-productive for mRNA export. So, what is their function? (5/x)
Most eukaryotes harbor two additional, less characterized TREX-2 like complexes: LENG8-PS and SAC3D1-PS, which interestingly differ in their subcellular localization. What is their function, and do they also act on UAP56? (4/x)
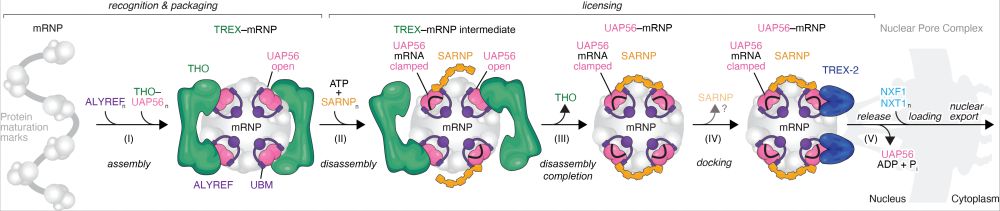
In the final step of mRNA export UAP56 enables docking of the mRNP (mRNA+bound proteins) to TREX-2, which is anchored at the nuclear pore. TREX-2 in turn removes UAP56 from the mRNA, enabling the release of the mRNP from the nuclear pore and its shuttling to the cytoplasm. (3/x)
We previously suggested a model for a core mRNA nuclear export pathway, which centers on the action of the RNA clampase UAP56: www.biorxiv.org/content/10.1... (2/x)