


Vanni Lab at UNIFR, Switzerland
@labvanni.bsky.social
Computational biophysics and other amenities...
"More thinking and less pipetting"
"More thinking and less pipetting"
Finally, @danialv.bsky.social used CG simulations to show that VPS13A can deliver lipids to an acceptor membrane that is bound at the C-terminal of VPS13A, and that XKR1 is proximal, but not continuous, to the delivery site. (6/7)
January 12, 2026 at 7:57 PM
Finally, @danialv.bsky.social used CG simulations to show that VPS13A can deliver lipids to an acceptor membrane that is bound at the C-terminal of VPS13A, and that XKR1 is proximal, but not continuous, to the delivery site. (6/7)
But how does VPS13A contact the lipid bilayer? Using a combination of coarse-grained (CG) MD simulation and in vitro flotation assay, we show that a disordered ATG2_C domain located in the C-terminal part of VPS13A is required for membrane binding. (5/7)

January 12, 2026 at 7:57 PM
But how does VPS13A contact the lipid bilayer? Using a combination of coarse-grained (CG) MD simulation and in vitro flotation assay, we show that a disordered ATG2_C domain located in the C-terminal part of VPS13A is required for membrane binding. (5/7)
The plasma membrane scramblase XKR1 directly interacts with the PH domain of VPS13A. All-atom MD simulations show that this interaction promotes a conformational change in XKR1 which exposes a hydrophilic groove, that acts as the scrambling region (4/7)

January 12, 2026 at 7:57 PM
The plasma membrane scramblase XKR1 directly interacts with the PH domain of VPS13A. All-atom MD simulations show that this interaction promotes a conformational change in XKR1 which exposes a hydrophilic groove, that acts as the scrambling region (4/7)
The cryo-EM data reveal that the VAB domain of VPS13A can adapt distinct conformations. These are likely to modulate the lipid transfer ability of VPS13A, and this mechanism is probably conserved in similar BLTPs. (3/7)

January 12, 2026 at 7:57 PM
The cryo-EM data reveal that the VAB domain of VPS13A can adapt distinct conformations. These are likely to modulate the lipid transfer ability of VPS13A, and this mechanism is probably conserved in similar BLTPs. (3/7)
The first step: at Yale, Bodan solved at near-atomic resolution the structure a prototypical BLTP, VSP13A, in complex with Calmodulin (CaM) and a plasma membrane scramblase, XKR1, demonstrating their physical interaction. (2/7)

January 12, 2026 at 7:57 PM
The first step: at Yale, Bodan solved at near-atomic resolution the structure a prototypical BLTP, VSP13A, in complex with Calmodulin (CaM) and a plasma membrane scramblase, XKR1, demonstrating their physical interaction. (2/7)
Happy to finally share the amazing results of our long-term collaboration with Karin Reinisch’s lab on how bridge lipid-transfer proteins (BLTPs) cooperate with partner proteins to orchestrate lipid delivery. A quick thread (1/7)
www.biorxiv.org/content/10.6...
www.biorxiv.org/content/10.6...
January 12, 2026 at 7:57 PM
Happy to finally share the amazing results of our long-term collaboration with Karin Reinisch’s lab on how bridge lipid-transfer proteins (BLTPs) cooperate with partner proteins to orchestrate lipid delivery. A quick thread (1/7)
www.biorxiv.org/content/10.6...
www.biorxiv.org/content/10.6...
Thanks to @snsf.ch, we are hiring two PhD students to investigate frontier topics in lipid biology using a combination of MD simulations and experiments. Preferred starting date: ASAP. Fribourg - and Switzerland - are quite nice, for both work and private life! Please spread the word!

October 13, 2025 at 2:17 PM
Thanks to @snsf.ch, we are hiring two PhD students to investigate frontier topics in lipid biology using a combination of MD simulations and experiments. Preferred starting date: ASAP. Fribourg - and Switzerland - are quite nice, for both work and private life! Please spread the word!
Happy to share the latest from the lab, led by Daniel Alvarez, in collaboration with @lizconibear.bsky.social. In this AA-MD tour-de-force, we delve deep into the mechanism and energetics of lipid uptake by bridge-like lipid transfer proteins, and we learn a few interesting things along the way...
August 7, 2025 at 8:45 AM
Happy to share the latest from the lab, led by Daniel Alvarez, in collaboration with @lizconibear.bsky.social. In this AA-MD tour-de-force, we delve deep into the mechanism and energetics of lipid uptake by bridge-like lipid transfer proteins, and we learn a few interesting things along the way...
We also modelled Brl1 in curved membranes (a vesicle mimicking the INM shape). Same result: Brl1/Brr6 oligomers stably interacting and promoting lipid mixing within the channel. (11/13)
July 23, 2025 at 11:53 AM
We also modelled Brl1 in curved membranes (a vesicle mimicking the INM shape). Same result: Brl1/Brr6 oligomers stably interacting and promoting lipid mixing within the channel. (11/13)
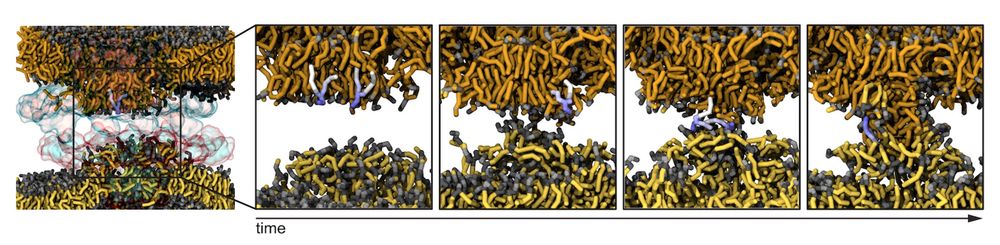
When observed closely, lipid mixing looks like classic hemifusion-like intermediates with exposed lipid-tails (but stabilized within protein rings here). (10/13)
July 23, 2025 at 11:51 AM
When observed closely, lipid mixing looks like classic hemifusion-like intermediates with exposed lipid-tails (but stabilized within protein rings here). (10/13)
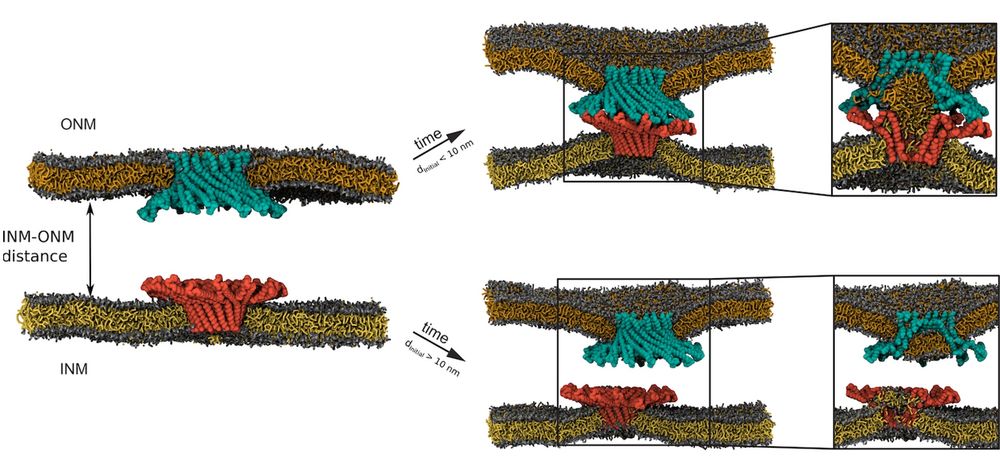
But here is another interesting part: interaction between Brl1 and Brr6 only happens when the membranes are < 10 nm apart. This is also very consistent with previous measurement of these distances in NPC assembly intermediates prior to fusion. (9/13)
July 23, 2025 at 11:51 AM
But here is another interesting part: interaction between Brl1 and Brr6 only happens when the membranes are < 10 nm apart. This is also very consistent with previous measurement of these distances in NPC assembly intermediates prior to fusion. (9/13)
We went back to simulations of opposed bilayers, but with protein oligomers in both of them. We observed a gradual decrease in membrane distances to the point of interaction between proteins (Brl1 and Brr6). This interaction results in lipid mixing in the central channel of the rings. (8/13)
July 23, 2025 at 11:50 AM
We went back to simulations of opposed bilayers, but with protein oligomers in both of them. We observed a gradual decrease in membrane distances to the point of interaction between proteins (Brl1 and Brr6). This interaction results in lipid mixing in the central channel of the rings. (8/13)
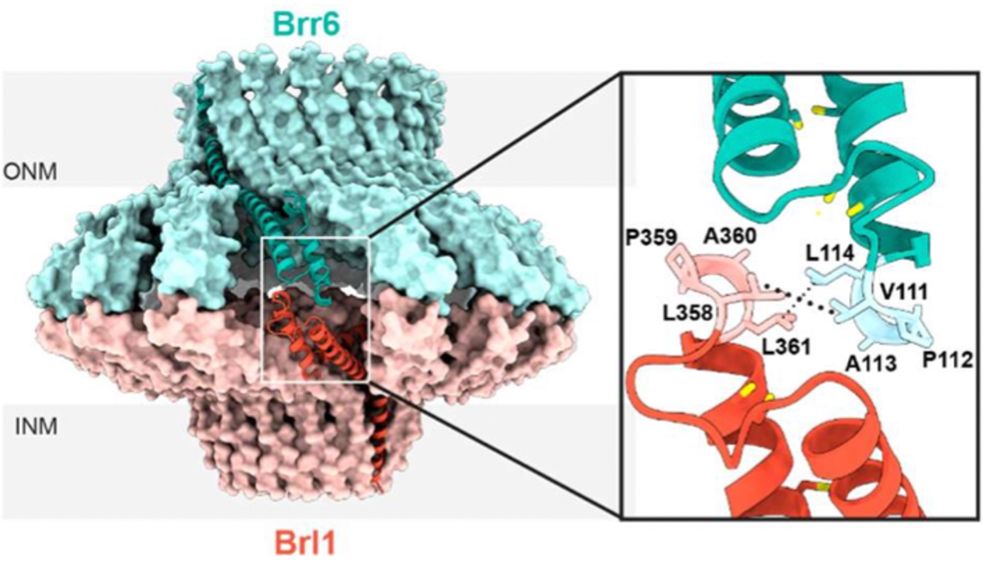
Given that both Brl1 and Brr6 are essential, and their interaction has been studied in earlier work, we modeled 16 copies of Brl1 together with 16 of Brr6. We got a 32-mer complex with two subunits of 16-mers of Brl1/Brr6 with a head-to-head interaction mediated by a hydrophobic loop motif. (6/13)
July 23, 2025 at 11:48 AM
Given that both Brl1 and Brr6 are essential, and their interaction has been studied in earlier work, we modeled 16 copies of Brl1 together with 16 of Brr6. We got a 32-mer complex with two subunits of 16-mers of Brl1/Brr6 with a head-to-head interaction mediated by a hydrophobic loop motif. (6/13)
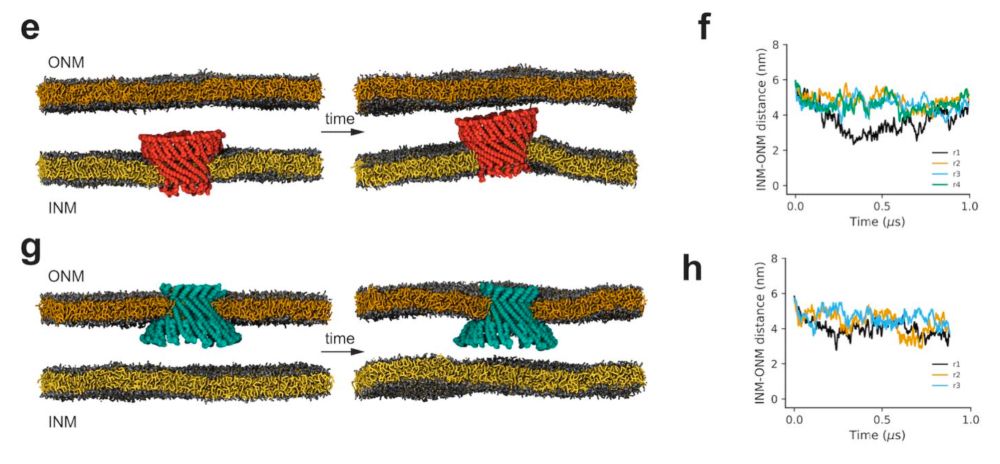
Our MD simulations of ring oligomers of Brl1/Brr6 showed pronounced lipid-remodeling in its vicinity. Could this lipid remodeling be enough to drive fusion of INM-ONM? The answer is NO, based on simulation of two opposed bilayers, with protein in only one of them. (5/13)
July 23, 2025 at 11:47 AM
Our MD simulations of ring oligomers of Brl1/Brr6 showed pronounced lipid-remodeling in its vicinity. Could this lipid remodeling be enough to drive fusion of INM-ONM? The answer is NO, based on simulation of two opposed bilayers, with protein in only one of them. (5/13)
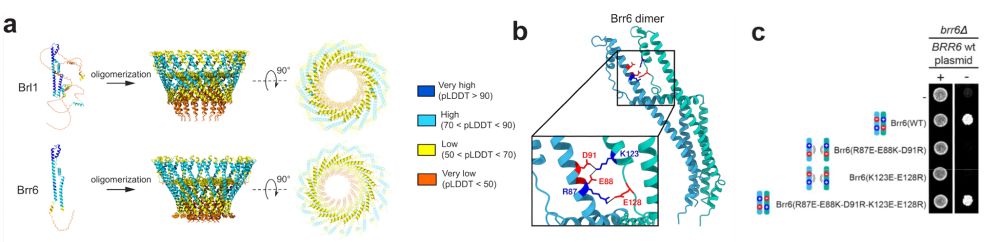
AF3 led us to find that both Brl1 and Brr6 tend to self-associate and form multimeric structures that become fully closed rings when 10 or more copies are present. By mutating charged residues at the dimer interface, we showed that this side-by-side oligomerization is critical. (4/13)
July 23, 2025 at 11:46 AM
AF3 led us to find that both Brl1 and Brr6 tend to self-associate and form multimeric structures that become fully closed rings when 10 or more copies are present. By mutating charged residues at the dimer interface, we showed that this side-by-side oligomerization is critical. (4/13)
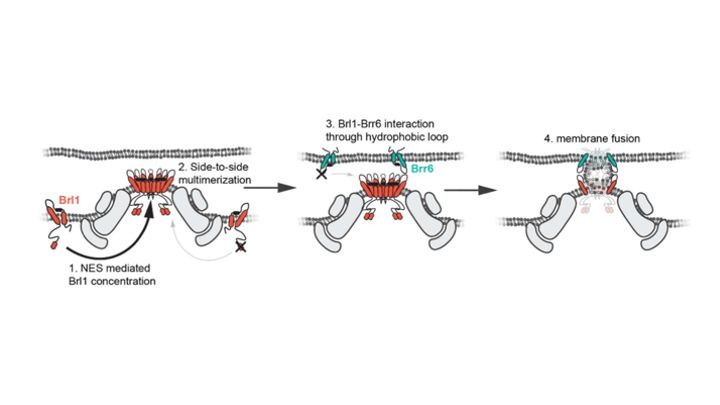
We found that Brl1 localizes to NPC assembly sites via an N-terminal nuclear export signal (NES). At that site, Brl1 and Brr6 oligomerize into rings, bridging membranes by interacting through conserved hydrophobic loops, promoting lipid mixing. Continue for more details! (3/13)
July 23, 2025 at 11:42 AM
We found that Brl1 localizes to NPC assembly sites via an N-terminal nuclear export signal (NES). At that site, Brl1 and Brr6 oligomerize into rings, bridging membranes by interacting through conserved hydrophobic loops, promoting lipid mixing. Continue for more details! (3/13)
Meet Brl1 and Brr6: two highly conserved transmembrane proteins in yeast, localized to the inner (INM) and outer nuclear membranes (ONM), respectively. Both are essential for NPC assembly, and despite their similarity, they are non-redundant. (2/13)
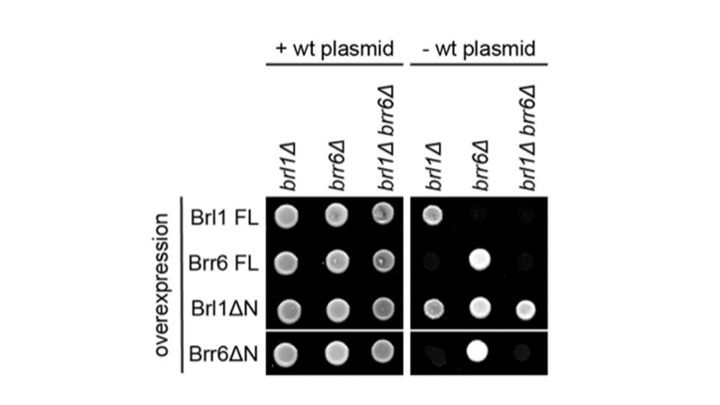
July 23, 2025 at 11:41 AM
Meet Brl1 and Brr6: two highly conserved transmembrane proteins in yeast, localized to the inner (INM) and outer nuclear membranes (ONM), respectively. Both are essential for NPC assembly, and despite their similarity, they are non-redundant. (2/13)
Happy to share the latest work from the lab, led by @mudgal17.bsky.social, in collaboration with the Weis lab @ethzurich.bsky.social.
How do nuclear membranes fuse during NPC assembly? We answer this question in our latest work, where we identify a new mechanism for membrane fusion… (1/13)
How do nuclear membranes fuse during NPC assembly? We answer this question in our latest work, where we identify a new mechanism for membrane fusion… (1/13)
July 23, 2025 at 11:40 AM
Happy to share the latest work from the lab, led by @mudgal17.bsky.social, in collaboration with the Weis lab @ethzurich.bsky.social.
How do nuclear membranes fuse during NPC assembly? We answer this question in our latest work, where we identify a new mechanism for membrane fusion… (1/13)
How do nuclear membranes fuse during NPC assembly? We answer this question in our latest work, where we identify a new mechanism for membrane fusion… (1/13)
Our lab is hiring 1 PhD and 1 Postdoc to study the mechanism of lipid transport using computational methods (Molecular Dynamics, AI). Fully funded by @snsf-ch.bsky.social for 4 years, with amazing collaborators and excellent life and working conditions. Apply by email. Please share and repost!
April 15, 2025 at 4:06 PM
Our lab is hiring 1 PhD and 1 Postdoc to study the mechanism of lipid transport using computational methods (Molecular Dynamics, AI). Fully funded by @snsf-ch.bsky.social for 4 years, with amazing collaborators and excellent life and working conditions. Apply by email. Please share and repost!
Join us in Heidelberg! Registration closes in 5 days!

November 8, 2024 at 1:06 PM
Join us in Heidelberg! Registration closes in 5 days!