


Takehiro A. Ozawa
@tkozawa.bsky.social
320 followers
740 following
32 posts
PhD candidate at CINVESTAV Unidad Irapuato 🇲🇽.
Interested in microalgae (e.g. Botryococcus braunii), algal biotechnology and plant energy management (i.e. #PlantTOR and #SnRK1 signaling pathways 🌱⚡️).
Posts
Media
Videos
Starter Packs
Reposted by Takehiro A. Ozawa



Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa



Reposted by Takehiro A. Ozawa

Reposted by Takehiro A. Ozawa




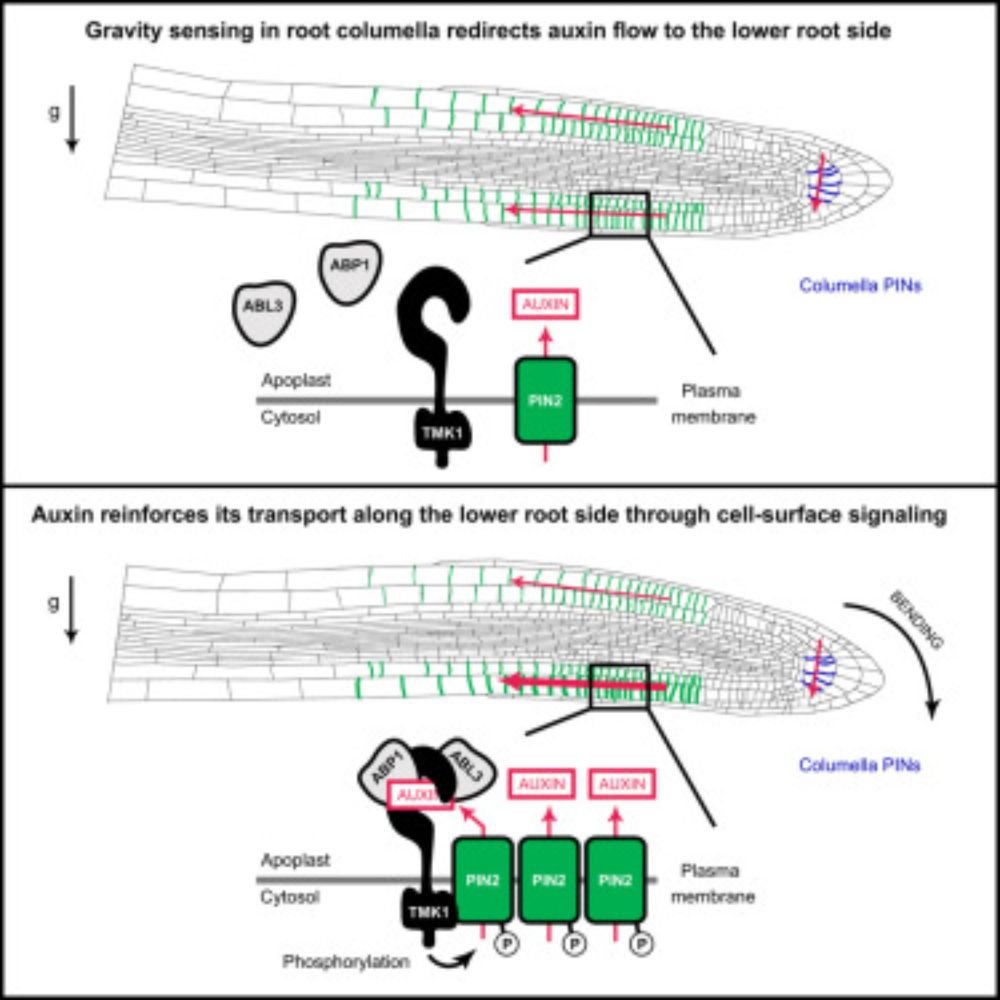
Reposted by Takehiro A. Ozawa





Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa




Reposted by Takehiro A. Ozawa






Reposted by Takehiro A. Ozawa


Reposted by Takehiro A. Ozawa
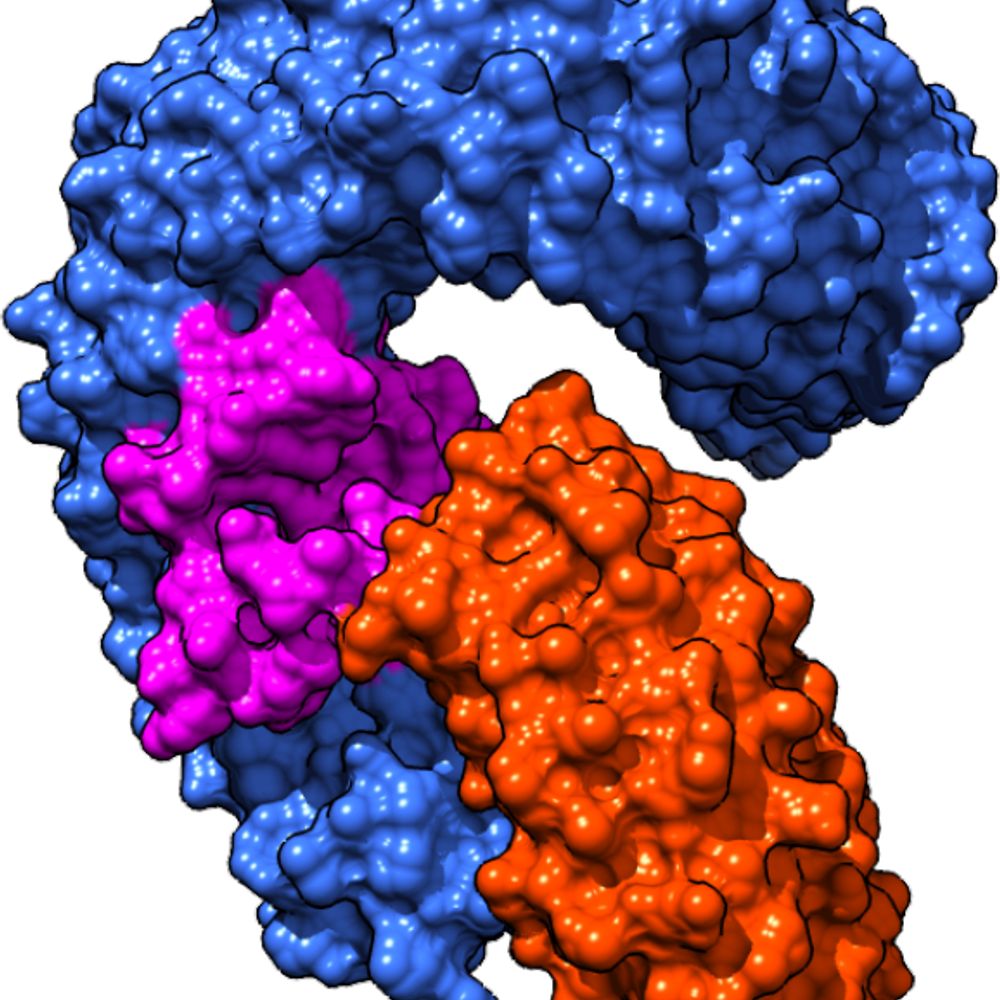

Reposted by Takehiro A. Ozawa




