



Alex Arzamasov
@arzamasovalex.bsky.social
Postdoc in Osterman lab at SBP Medical Discovery Institute.
Interested in the functional annotation of genomic and metagenomic data, carbohydrate metabolism, #bifidobacteria
Interested in the functional annotation of genomic and metagenomic data, carbohydrate metabolism, #bifidobacteria
Pinned
Alex Arzamasov
@arzamasovalex.bsky.social
· Sep 30
It’s been a bit over 2 months since the main results of my PhD work on reconstructing carbohydrate utilization pathways in human bifidobacteria were finally published; so I guess it’s a good time to update the previous thread (1/7) www.nature.com/articles/s41...
Reposted by Alex Arzamasov

Have a look if you are interested in how the team at @cultivarium.bsky.social is finding electroporation protocols for new microbes: both screening lots of conditions on a custom built electroporator concurrently, and cyclical iteration using bayesian optimization
www.biorxiv.org/content/10.1...
www.biorxiv.org/content/10.1...

Active learning guides automated discovery of DNA delivery via electroporation for non-model microbes
Delivery of recombinant DNA is foundational for understanding and engineering a target organism. Electroporation can be applied to any cell type, yet identification of a working protocol for new organ...
www.biorxiv.org
November 22, 2025 at 12:23 AM
Have a look if you are interested in how the team at @cultivarium.bsky.social is finding electroporation protocols for new microbes: both screening lots of conditions on a custom built electroporator concurrently, and cyclical iteration using bayesian optimization
www.biorxiv.org/content/10.1...
www.biorxiv.org/content/10.1...
Overall, given that FOS are often included in infant formulas and used widely as prebiotics, it’s important to understand the mechanisms by which microbes utilize these oligosaccharides of different chain lengths
November 16, 2025 at 1:48 AM
Overall, given that FOS are often included in infant formulas and used widely as prebiotics, it’s important to understand the mechanisms by which microbes utilize these oligosaccharides of different chain lengths
We observed a similar pattern in Bifidobacterium: only strains carrying a specific ABC transporter could metabolize long-chain FOS, while utilization of short-chain FOS was broadly conserved
www.nature.com/articles/s41...
www.nature.com/articles/s41...

Integrative genomic reconstruction reveals heterogeneity in carbohydrate utilization across human gut bifidobacteria - Nature Microbiology
A comprehensive genomic analysis reveals species- and strain-level heterogeneity in carbohydrate utilization potential across bifidobacteria of human origin.
www.nature.com
November 16, 2025 at 1:48 AM
We observed a similar pattern in Bifidobacterium: only strains carrying a specific ABC transporter could metabolize long-chain FOS, while utilization of short-chain FOS was broadly conserved
www.nature.com/articles/s41...
www.nature.com/articles/s41...
Very cool print demonstrating how variation in at TonB-dependent transporter repertoires shapes the ability of Bacteroides and Phocaeicola species to utilize fructooligosaccharides (FOS) of different chain lengths
Niche partitioning by resource size in the gut microbiome https://www.biorxiv.org/content/10.1101/2025.11.13.688124v1
November 16, 2025 at 1:48 AM
Very cool print demonstrating how variation in at TonB-dependent transporter repertoires shapes the ability of Bacteroides and Phocaeicola species to utilize fructooligosaccharides (FOS) of different chain lengths
Some cool data on bifidobacteria and their relatives in honeybees!
Happy to announce that our comparative metagenomic analysis of the gut microbiota of five honeybee species - spearheaded by @aiswarya.bsky.social - is finally published in a peer-reviewed journal! rdcu.be/eKMCs @fbm-unil.bsky.social @dmf-unil.bsky.social 🧵👇

Evolution of gut microbiota across honeybee species revealed by comparative metagenomics
Nature Communications - Here, the authors perform metagenomic analysis of honeybee gut microbes, uncovering many previously unidentified species and host specific differences in composition and...
rdcu.be
October 14, 2025 at 9:48 PM
Some cool data on bifidobacteria and their relatives in honeybees!
Reposted by Alex Arzamasov

Happy to announce that our comparative metagenomic analysis of the gut microbiota of five honeybee species - spearheaded by @aiswarya.bsky.social - is finally published in a peer-reviewed journal! rdcu.be/eKMCs @fbm-unil.bsky.social @dmf-unil.bsky.social 🧵👇
Evolution of gut microbiota across honeybee species revealed by comparative metagenomics
Nature Communications - Here, the authors perform metagenomic analysis of honeybee gut microbes, uncovering many previously unidentified species and host specific differences in composition and...
rdcu.be
October 14, 2025 at 7:04 PM
Happy to announce that our comparative metagenomic analysis of the gut microbiota of five honeybee species - spearheaded by @aiswarya.bsky.social - is finally published in a peer-reviewed journal! rdcu.be/eKMCs @fbm-unil.bsky.social @dmf-unil.bsky.social 🧵👇
Genomic studies in bifidobacteria often focus only on CAZymes. IMHO, in Gram-positives, transporters are the true gatekeepers of glycan metabolism. We need to prioritize improving their functional annotations and include them in metabolic reconstructions (7/7)
September 30, 2025 at 9:01 AM
Genomic studies in bifidobacteria often focus only on CAZymes. IMHO, in Gram-positives, transporters are the true gatekeepers of glycan metabolism. We need to prioritize improving their functional annotations and include them in metabolic reconstructions (7/7)
We demonstrated that phylogenetically closely related strains can exhibit substantial differences in HMO utilization, which is driven by subtle variations in HMO transporter genes. Species/subspecies names alone don’t tell the full story; one needs to look at gene content in individual strains(6/7)

September 30, 2025 at 9:01 AM
We demonstrated that phylogenetically closely related strains can exhibit substantial differences in HMO utilization, which is driven by subtle variations in HMO transporter genes. Species/subspecies names alone don’t tell the full story; one needs to look at gene content in individual strains(6/7)
Among the new pathways we uncovered was a xyloglucan degradation pathway that was present in rare B. catenulatum subsp. kashiwanohense strains and conserved in B. dentium and B. tsurumiense (5/7)

September 30, 2025 at 9:01 AM
Among the new pathways we uncovered was a xyloglucan degradation pathway that was present in rare B. catenulatum subsp. kashiwanohense strains and conserved in B. dentium and B. tsurumiense (5/7)
We validated phenotypic predictions for 30 bifidobacterial strains, achieving 94% accuracy. For example, we confirmed the unique ability of the new B. longum clade to grow on starch and pullulan, and described an unconventional B. adolescentis strain that can use 2’-fucosyllactose (4/7)

September 30, 2025 at 9:01 AM
We validated phenotypic predictions for 30 bifidobacterial strains, achieving 94% accuracy. For example, we confirmed the unique ability of the new B. longum clade to grow on starch and pullulan, and described an unconventional B. adolescentis strain that can use 2’-fucosyllactose (4/7)
Our analysis revealed notable inter- and intra-species variability. Among notable findings was a new Bifidobacterium longum clade harboring pathways for starch, pullulan, and difructose dianhydride metabolism but lacking pathways for LNB/GNB, N-glycan, and human milk oligosaccharide utilization(3/7)

September 30, 2025 at 9:01 AM
Our analysis revealed notable inter- and intra-species variability. Among notable findings was a new Bifidobacterium longum clade harboring pathways for starch, pullulan, and difructose dianhydride metabolism but lacking pathways for LNB/GNB, N-glycan, and human milk oligosaccharide utilization(3/7)
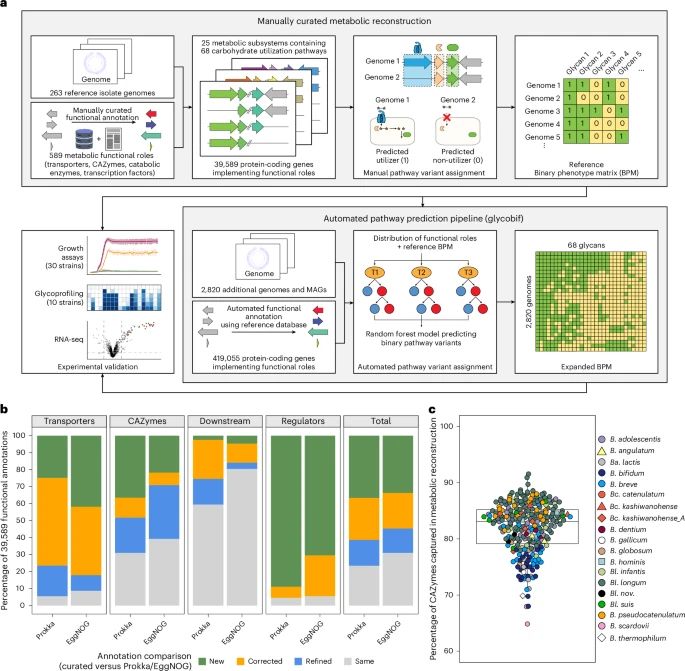
We reconstructed 68 glycan utilization pathways encoded in 3,083 bif genomes by looking at the distribution of 589 curated metabolic functions (transporters, CAZymes, etc). Several years of manual curation greatly improved the quality of functional gene annotations (>90% for transporters!) (2/7)

September 30, 2025 at 9:01 AM
We reconstructed 68 glycan utilization pathways encoded in 3,083 bif genomes by looking at the distribution of 589 curated metabolic functions (transporters, CAZymes, etc). Several years of manual curation greatly improved the quality of functional gene annotations (>90% for transporters!) (2/7)
It’s been a bit over 2 months since the main results of my PhD work on reconstructing carbohydrate utilization pathways in human bifidobacteria were finally published; so I guess it’s a good time to update the previous thread (1/7) www.nature.com/articles/s41...
September 30, 2025 at 9:01 AM
It’s been a bit over 2 months since the main results of my PhD work on reconstructing carbohydrate utilization pathways in human bifidobacteria were finally published; so I guess it’s a good time to update the previous thread (1/7) www.nature.com/articles/s41...
Among the new pathways we uncovered was a xyloglucan degradation pathway that was present in rare B. catenulatum subsp. kashiwanohense strains and conserved in B. dentium and B. tsurumiense (5/7)

September 30, 2025 at 8:49 AM
Among the new pathways we uncovered was a xyloglucan degradation pathway that was present in rare B. catenulatum subsp. kashiwanohense strains and conserved in B. dentium and B. tsurumiense (5/7)
We validated phenotypic predictions for 30 bifidobacterial strains, achieving 94% accuracy. For example, we confirmed the unique ability of the new B. longum clade to grow on starch and pullulan, and described a B. adolescentis strain that can use 2’-fucosyllactose (4/7)

September 30, 2025 at 8:49 AM
We validated phenotypic predictions for 30 bifidobacterial strains, achieving 94% accuracy. For example, we confirmed the unique ability of the new B. longum clade to grow on starch and pullulan, and described a B. adolescentis strain that can use 2’-fucosyllactose (4/7)
Our analysis revealed notable inter- and intra-species variability. Among notable findings was a new Bifidobacterium longum clade harboring pathways for starch, pullulan, and difructose dianhydride metabolism but lacking pathways for LNB/GNB, N-glycan, and human milk oligosaccharide utilization(3/7)

September 30, 2025 at 8:49 AM
Our analysis revealed notable inter- and intra-species variability. Among notable findings was a new Bifidobacterium longum clade harboring pathways for starch, pullulan, and difructose dianhydride metabolism but lacking pathways for LNB/GNB, N-glycan, and human milk oligosaccharide utilization(3/7)
We reconstructed 68 glycan utilization pathways encoded in 3,083 bif genomes by looking at the distribution of 589 curated metabolic functions (transporters, CAZymes, etc). Several years of manual curation greatly improved the quality of functional gene annotations (>90% for transporters!) (2/7)
September 30, 2025 at 8:49 AM
We reconstructed 68 glycan utilization pathways encoded in 3,083 bif genomes by looking at the distribution of 589 curated metabolic functions (transporters, CAZymes, etc). Several years of manual curation greatly improved the quality of functional gene annotations (>90% for transporters!) (2/7)
Reposted by Alex Arzamasov
Excited to share our @cp-cellhostmicrobe.bsky.social led by Magda showing how Bif has co-evolved with different animal hosts 🐒🐭🐷🐦
Key takeaways:
🔹 Host ancestry + diet shape Bif evolution
🔹 Mammals enriched for carb-busting enzymes
🔹 Untapped diversity in non-human hosts = new probiotic potential
Key takeaways:
🔹 Host ancestry + diet shape Bif evolution
🔹 Mammals enriched for carb-busting enzymes
🔹 Untapped diversity in non-human hosts = new probiotic potential

Bifido adaptations across hosts
@halllab.bsky.social survey insects, reptiles, birds & mammals to uncover how Bifidobacterium adapts to hosts. Bifidobacterium & host exhibit strong co-phylogenetic associations, driven by vertical transmission & dietary selection
www.cell.com/cell-host-mi...
@halllab.bsky.social survey insects, reptiles, birds & mammals to uncover how Bifidobacterium adapts to hosts. Bifidobacterium & host exhibit strong co-phylogenetic associations, driven by vertical transmission & dietary selection
www.cell.com/cell-host-mi...

Host-specific microbiome and genomic signatures in Bifidobacterium reveal co-evolutionary and functional adaptations across diverse animal hosts
Bifidobacterium are beneficial members of the microbiota across animal hosts. Kujawska
et al. demonstrate that bifidobacteria have likely co-evolved more closely with mammals,
particularly primates, a...
www.cell.com
September 11, 2025 at 2:49 PM
Excited to share our @cp-cellhostmicrobe.bsky.social led by Magda showing how Bif has co-evolved with different animal hosts 🐒🐭🐷🐦
Key takeaways:
🔹 Host ancestry + diet shape Bif evolution
🔹 Mammals enriched for carb-busting enzymes
🔹 Untapped diversity in non-human hosts = new probiotic potential
Key takeaways:
🔹 Host ancestry + diet shape Bif evolution
🔹 Mammals enriched for carb-busting enzymes
🔹 Untapped diversity in non-human hosts = new probiotic potential
Reposted by Alex Arzamasov

Our study developing a skin metatranscriptomics protocol is now out in @natbiotech.nature.com!
We finally have the ability to study microbial activity on skin and identify key functional genes playing a role in diseases.
Amazing team of Chia Minghao and Amanda Ng 👏
nature.com/articles/s41...
We finally have the ability to study microbial activity on skin and identify key functional genes playing a role in diseases.
Amazing team of Chia Minghao and Amanda Ng 👏
nature.com/articles/s41...
August 30, 2025 at 2:17 AM
Our study developing a skin metatranscriptomics protocol is now out in @natbiotech.nature.com!
We finally have the ability to study microbial activity on skin and identify key functional genes playing a role in diseases.
Amazing team of Chia Minghao and Amanda Ng 👏
nature.com/articles/s41...
We finally have the ability to study microbial activity on skin and identify key functional genes playing a role in diseases.
Amazing team of Chia Minghao and Amanda Ng 👏
nature.com/articles/s41...
Very nice paper that deciphers the transport mechanisms of fucosylated lacto-N-biose in Bifidobacterium longum subsp. infantis
journals.asm.org/doi/full/10....
journals.asm.org/doi/full/10....

Uptake of fucosylated type I human milk oligosaccharide blocks by Bifidobacterium longum subsp. infantis | mBio
The assembly of the gut microbiota in early life is critical to the health trajectory
of human hosts. Breast feeding selects for a Bifidobacterium-rich community, adapted to efficiently utilize human ...
journals.asm.org
July 26, 2025 at 4:19 AM
Very nice paper that deciphers the transport mechanisms of fucosylated lacto-N-biose in Bifidobacterium longum subsp. infantis
journals.asm.org/doi/full/10....
journals.asm.org/doi/full/10....
Reposted by Alex Arzamasov

High-throughput single-cell isolation of Bifidobacterium strains from the gut microbiome https://www.biorxiv.org/content/10.1101/2025.07.23.666462v1
July 25, 2025 at 4:17 AM
High-throughput single-cell isolation of Bifidobacterium strains from the gut microbiome https://www.biorxiv.org/content/10.1101/2025.07.23.666462v1
Thanks! Also, maybe of interest, I think PSORTb and the new tool DeepLocPro perform better for bifido proteins than signalP
academic.oup.com/bioinformati...
academic.oup.com/bioinformati...

Predicting the subcellular location of prokaryotic proteins with DeepLocPro
AbstractMotivation. Protein subcellular location prediction is a widely explored task in bioinformatics because of its importance in proteomics research. W
academic.oup.com
July 19, 2025 at 2:13 AM
Thanks! Also, maybe of interest, I think PSORTb and the new tool DeepLocPro perform better for bifido proteins than signalP
academic.oup.com/bioinformati...
academic.oup.com/bioinformati...
4) The GH136 lacto-N-biosidase from B. longum that you cite does not release LacNAc from Lacto-N-neotetraose (Fig. 4b from Sakurama et al). The B. bifidum GH136 is not identical to the B. longum one, but I haven't seen papers describing activity on LNnT (only LNT)
www.jbc.org/article/S002...
www.jbc.org/article/S002...

Lacto-N-biosidase Encoded by a Novel Gene of Bifidobacterium longum Subspecies longum Shows Unique Substrate Specificity and Requires a Designated Chaperone for Its Active Expression *
Infant gut-associated bifidobacteria possess species-specific enzymatic sets to assimilate
human milk oligosaccharides, and lacto-N-biosidase (LNBase) is a key enzyme that degrades
lacto-N-tetraose (G...
www.jbc.org
July 18, 2025 at 4:47 PM
4) The GH136 lacto-N-biosidase from B. longum that you cite does not release LacNAc from Lacto-N-neotetraose (Fig. 4b from Sakurama et al). The B. bifidum GH136 is not identical to the B. longum one, but I haven't seen papers describing activity on LNnT (only LNT)
www.jbc.org/article/S002...
www.jbc.org/article/S002...
3) Additionally, B. bifidum possesses an ortholog of an ABC transporter that can uptake GNB (GltABC), so it would make sense for the GH101 enzyme to be extracellular, so the released disaccharide is then imported.
www.jbc.org/article/S002...
www.jbc.org/article/S002...

Structural and Thermodynamic Analyses of Solute-binding Protein from Bifidobacterium longum Specific for Core 1 Disaccharide and Lacto-N-biose I *
Recently, a gene cluster involving a phosphorylase specific for lacto-N-biose I (LNB;
Galβ1–3GlcNAc) and galacto-N-biose (GNB; Galβ1–3GalNAc) has been found in Bifidobacterium
longum. We showed that t...
www.jbc.org
July 18, 2025 at 4:47 PM
3) Additionally, B. bifidum possesses an ortholog of an ABC transporter that can uptake GNB (GltABC), so it would make sense for the GH101 enzyme to be extracellular, so the released disaccharide is then imported.
www.jbc.org/article/S002...
www.jbc.org/article/S002...
2) Your predictions about the localization of some other GHs (like GH89 and GH101) also differ from previously published literature. For example, the GH89 was described as a membrane-anchored extracellular enzyme before.
link.springer.com/article/10.1...
link.springer.com/article/10.1...

α-N-Acetylglucosaminidase from Bifidobacterium bifidum specifically hydrolyzes α-linked N-acetylglucosamine at nonreducing terminus of O-glycan on gastric mucin - Applied Microbiology and Biotechnolog...
α-Linked N-acetylglucosamine is one of the major glyco-epitopes in O-glycan of gastroduodenal mucin. Here, we identified glycoside hydrolase (GH) family 89 α-N-acetylglucosaminidase, termed AgnB, from...
link.springer.com
July 18, 2025 at 4:47 PM
2) Your predictions about the localization of some other GHs (like GH89 and GH101) also differ from previously published literature. For example, the GH89 was described as a membrane-anchored extracellular enzyme before.
link.springer.com/article/10.1...
link.springer.com/article/10.1...