
Axel Delamarre
@axeldelamarre.bsky.social
1.4K followers
1.5K following
19 posts
I study Chromatin structure, organization and function.
Postdoc in the Whitehouse lab, MSKCC, NYC.
Former PhD in the Pasero lab, IGH, France.
Posts
Media
Videos
Starter Packs
Pinned

Axel Delamarre
@axeldelamarre.bsky.social
· Jul 17




Reposted by Axel Delamarre
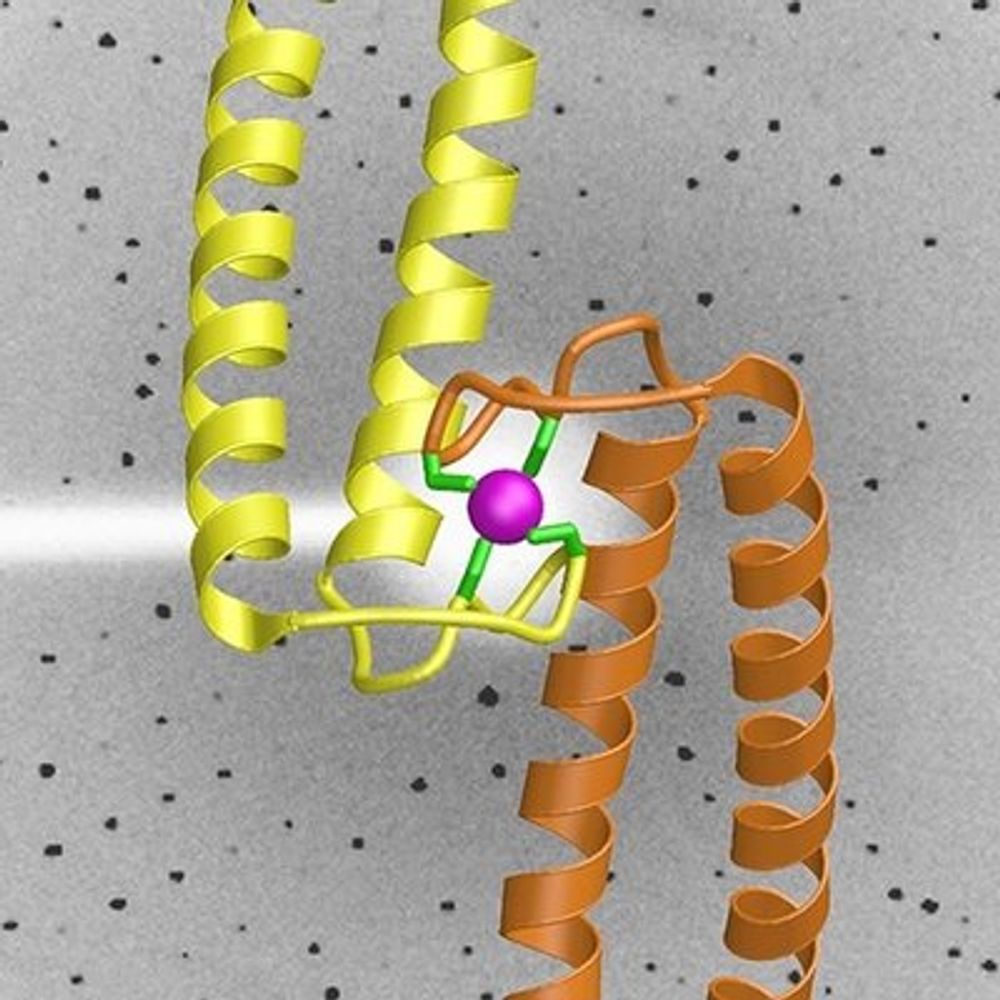


Reposted by Axel Delamarre

Reposted by Axel Delamarre


Reposted by Axel Delamarre

Mathew Jones
@jonesmjk.bsky.social
· Sep 1

A high-resolution, nanopore-based artificial intelligence assay for DNA replication stress in human cancer cells
Nature Communications - Determining how replication forks move across the human genome is critical for the effective use of agents that target replication stress. Here, the authors present...
rdcu.be
Reposted by Axel Delamarre


Reposted by Axel Delamarre





Reposted by Axel Delamarre


Reposted by Axel Delamarre

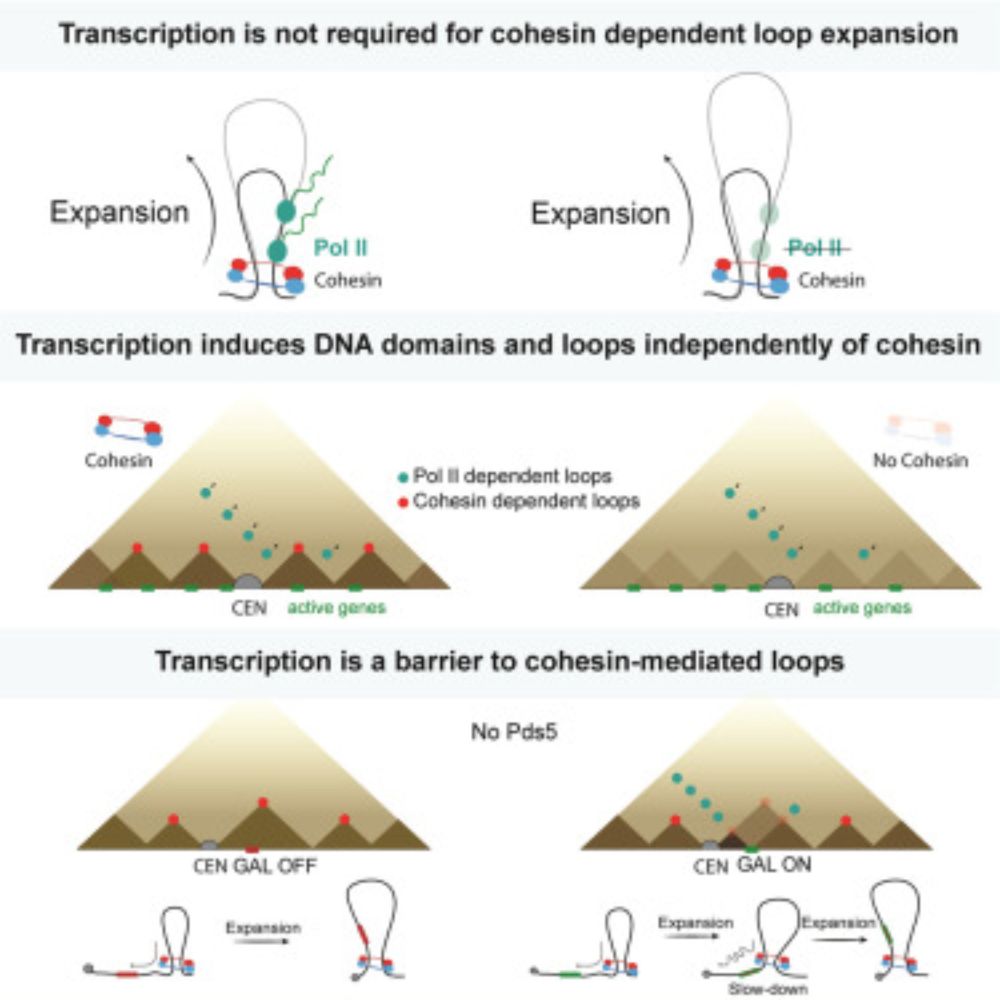
Reposted by Axel Delamarre
NealeLab
@labneale.bsky.social
· Aug 10

Condensin and topoisomerases cooperate to relieve topological stress at stalled replication forks
Resolving complex topological structures at replication forks is vital for successful DNA replication, but the mechanisms are little understood. Evidence from diverse eukaryotes suggests that condensi...
biorxiv.org
Reposted by Axel Delamarre


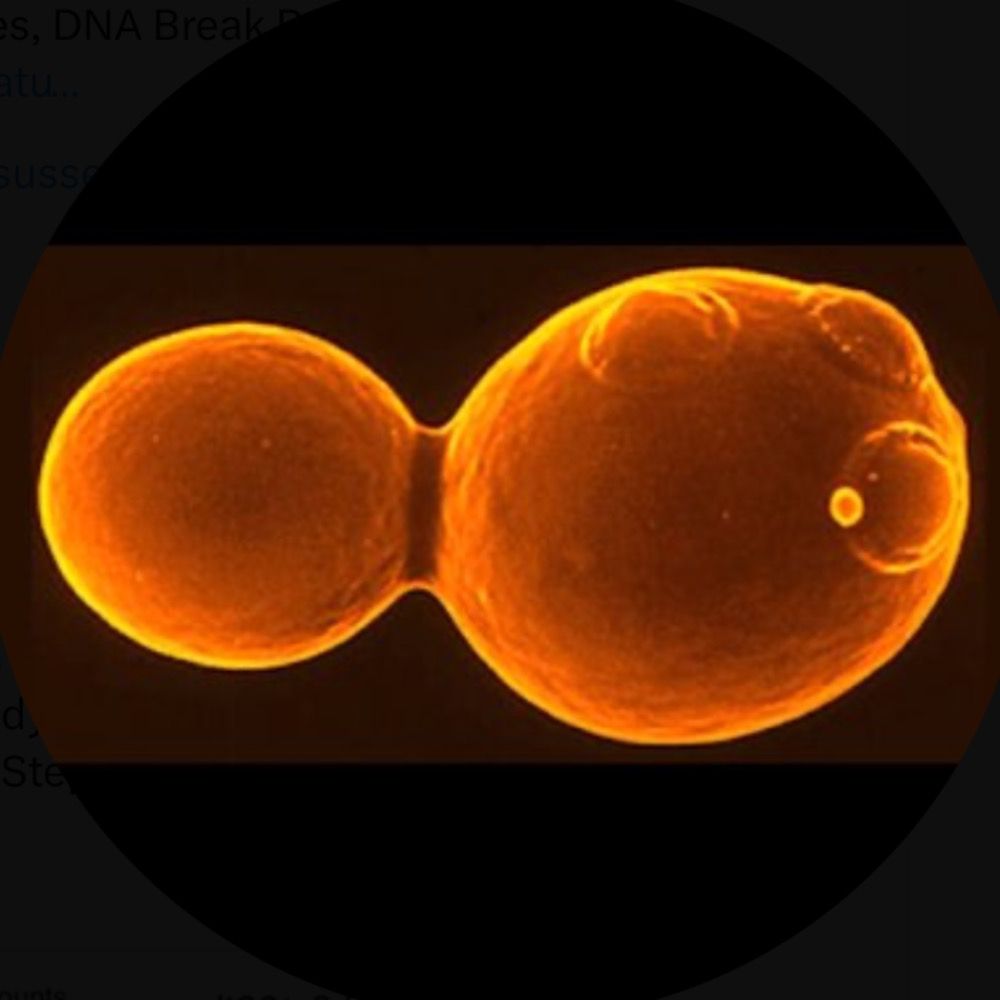
Axel Delamarre
@axeldelamarre.bsky.social
· Jul 17

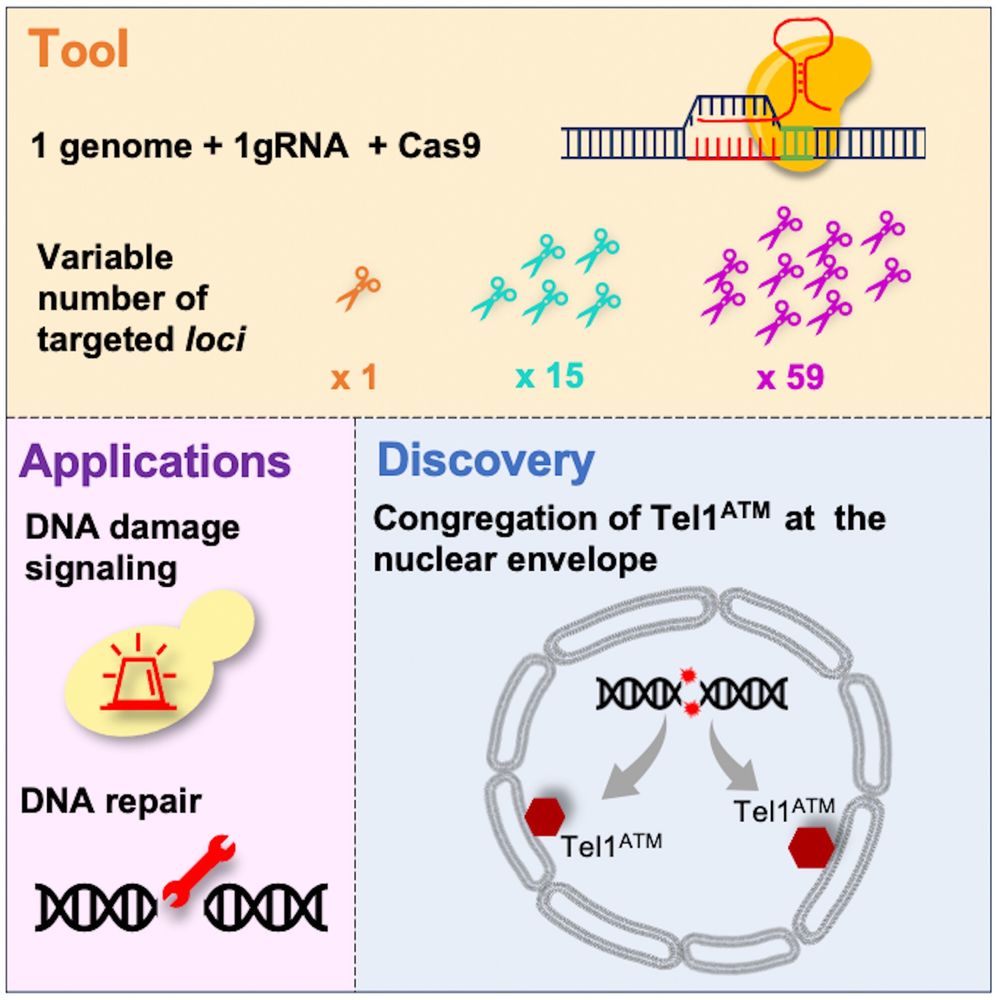
Axel Delamarre
@axeldelamarre.bsky.social
· May 30

Chromatin architecture mapping by multiplex proximity tagging
Chromatin plays a pivotal role in genome expression, maintenance, and replication. To better understand chromatin organization, we developed a novel proximity-tagging method which assigns unique DNA b...
www.biorxiv.org








Axel Delamarre
@axeldelamarre.bsky.social
· May 30